
In Darwinian terms, sex is good for the community, bad for the individual. If this statement doesn’t make sense to you, perhaps you are thinking that sex is a part of reproduction, and reproduction is the very definition of Darwinian fitness.
But sex is the sharing of genes. Reproduction is the process of creating new individuals. There is no logically necessary link between the two. In most higher organisms, evolution has linked gene-sharing to reproduction as a way to force individuals to share genes.
In most higher organisms, evolution has linked gene-sharing to
reproduction as a way to force individuals to share genes.
Which brings me back to the opening statement: By neo-Darwinian standards, the ideal fitness strategy would be to clone oneself endlessly. But this leads to a collapse of genetic diversity and a fragile population. Sharing genes is essential for a healthy population, but it is only a distraction in pursuit of “individual fitness”. If I clone myself, 100% of my genes are transferred to the next generation. If I reproduce sexually, only 50% of my genes get into the next generation. And if I have a special combination of genes that make me extra-fit, that combination is sure to be broken up in the mixing process that is sex.
The strongest clue we have that evolution cares about populations and not just individuals is the fact that, in most multi-celled life forms, sex is a prerequisite to reproduction. Evolution has arranged it so that sharing genes is mandated. You want to reproduce? You’re going to have to mix your genes with someone else’s.
Seen in this way, obligate sex is a grand communist conspiracy to hold back the strongest individuals and force them to contribute to the community, rather than to “get ahead” on their own.
Most people I know don’t need a lot of motivators in order to have sex. Biology has supplied the motivation. At one level, evolution has given us a powerful sex drive. At a deeper level, evolution has made sure that gene sharing is a prerequisite to reproduction, so that the drive to reproduce is transformed into a drive to copulate.
Ciliates and other protists
Single-celled organisms are of three types: Bacteria, archaea, and protists. Small, small, and large. Simple, simple, and complex.
Protists are the large, complicated protozoa from which multi-celled life emerged. The single cell is a million times larger than a bacterial cell and far more complex and structured; but still it’s a single cell.
Paramecia are ciliates. They are an advanced form of protists, capable of changing shape, moving about, and sophisticated goal-directed behaviors. Paramecia can learn and remember, though they obviously have no brains or even nervous systems. [another ref] No one knows how they acquire information or store it. A fascinating topic, but not for today.
This is lacrymaria, an advanced ciliate. It is a single cell that shapes itself to form a head and neck that can hunt for food.
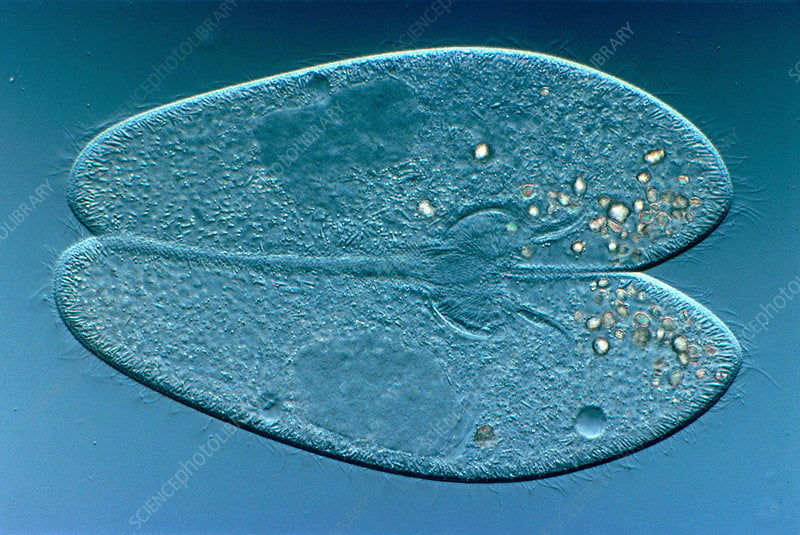
In paramecia, reproduction and sex are entirely separate functions. Reproduction is clonal. The paramecium splits in half, its DNA is copied with half going to each daughter cell. Sex is via a process known as conjugation. Two paramecia sidle up to each other, and their cells merge, their chromosomes pair up and some genes cross over (as in sex for higher organisms). Two paramecia separate, but they are not the same two that came together a few hours earlier. Each individual is “half me and half you”. Conjugation can require many hours in some species. Sharing genes is much slower and, presumably, more costly than reproduction.
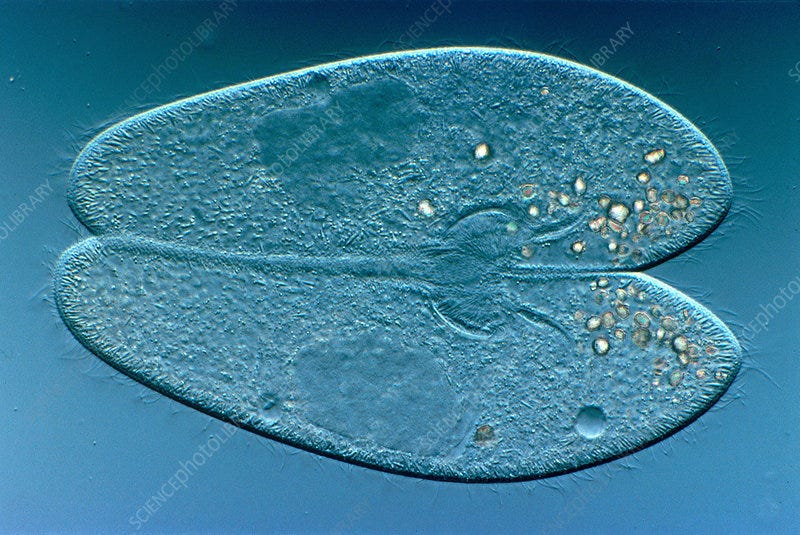
Conjugating paramecia, with merged micronuclei and separate macronuclei visible.
There are boy paramecia and girl paramecia. Two sexes. Other ciliates have multiple sexes with complicated rules about who can mate with whom. (The Tinder app for ciliates is way over budget and past its development deadline.)
Protist genetics
You and I each have one single genome, a set of 23 pairs of chromosome that sits in the nucleus of every cell in our bodies. Complicated chemical tags are attached to the chromosomes that determine which genes are active when and where. The DNA in each cell is actively being transcribed all the time, and this transcription controls the cell’s metabolism.
Ciliates have a different system. Each cell has a micronucleus containing a pristine copy of the DNA, and a macronucleus, with multiple copies of each gene in a DNA loop called a plasmid. The macronucleus is the workhorse, from which all gene products are transcribed, while the micronucleus retains a single archival copy of each chromosome.
When a paramecium splits in two, the micronucleus makes a copy of every chromosome, so there is a fresh new micronucleus for each daughter cell. The macronucleus, however, just splits in two, in a process called amitosis. Each daughter cell gets half of the macronucleus, containing an arbitrary selection of half the plasmids.
Remember that the macronucleus contains many copies of each gene, so usually the arbitrary split results in at least one copy of each gene in each half. The macronucleus regrows itself as the daughter cell grows and matures. Plasmids in the macronucleus are always copying themselves, creating copies of copies, but the archival copy remains in the micronucleus, where it is protected from chemical disturbances.
Over hundreds of cell divisions, the macronucleus becomes dysfunctional. The cell languishes and cannot continue growing and reproducing. This is cell senescence, ciliate style. (It does not have to do with telomere length, which is a trigger of cell senescence in multicelled organisms.)
The only cure for this “disease of old age” is to create a new macronucleus, using the archival chromosomes in the micronucleus.
Any self-respecting, rugged individual paramecium would just spit out its old macronucleus and create a new one. But they have been tamed. There are no rugged individuals left, and they all have to live by the rule. The rule says that you don’t get to make a new macronucleus — even if your life depends on it — unless you share your genes.
Renewal is only available to the cell pursuant to conjugation. A new macronucleus is only created after gene exchange.
How did senescence in Ciliates evolve?
The process described above is a kind of senescence for ciliates. After one or two hundred cell divisions, the cell line loses its ability to reproduce. The macronucleus poops out, for reasons that are not fully understood. Perhaps it is mutation or DNA damage. Perhaps the different copies of copies of copies of plasmids in the macronucleus become unbalanced, with too many copies of some plasmids and a shortage of others. In any case, the cell needs a new macronucleus.
The interesting thing is that the cell is perfectly capable of creating a new macronucleus from the archival copy of DNA stored in its micronucleus, but it does not do this.
Evolution has decreed that the creation of a new macronucleus from the micronucleus can only occur subsequent to conjugation. In other words, aging and death are built into the life plan for paramecia, and sharing genes is the only way to get a new start.
Contrast this to the situation for mammals like us. In both ciliates and mammals, sharing of genes is compulsory. But in mammals, sharing of genes is enforced by making sex a prerequisite for reproduction; while in ciliates, it is aging that enforces gene-sharing. The paramecia offspring will eventually get old and die if they don’t conjugate, and conjugation resets the clock for a new lease on life.
Notice that the cell that gets a new lease on life is not the fabulously successful cell that had created hundreds of generations of copies of itself. That genome has reached a dead end. The cell that gets to start over with a new micronucleus is a hybrid, with half its genes from each of two cell lines.
“Aha,” you say, “I’m going to beat the system by cloning, then conjugating with my twin. That way I can hold on to this very special combinations that is me me ME.”
Not so fast. There are at least two different sex types, and your twin is the same as your type. You can’t inbreed. It’s against the rules.
You can clone yourself for awhile, but your clonal lineage is a dead end. Only the community survives long-term, not the individual.
How did sex first become mandatory?
What was the evolutionary mechanism by which aging was imposed on paramecia, and gene-sharing was the only means by which they could escape the grim reaper? AFAIK, I have been the only one to ask this question.
(There has been lots of academic attention to the evolution of sex, which is long recognized as an evolutionary mystery. But little or none to this closely-related mechanism from hundreds of millions of years earlier.)
I created a computer model a few years ago, but was unable to publish it at the time. The peer review system worked as it should, and reviewers pointed out to me that I had misunderstood the dynamics of senescence and reproduction in ciliates. I have recently rewritten the ms to reflect the understanding that I describe above, and yesterday resubmitted the ms for publication. In the meantime, the preprint is on ResearchGate and on BioRxiv.
My ms actually includes two computer models, reflecting the mysterious nature of the phenomenon I’m modeling. I don’t claim to explain how senescence evolved, only to offer scenarios as a start to exploring this important transition in evolutionary history.
Population diversity is important for two principal reasons.
- Diversity is the fuel for evolutionary change. A homogeneous population can only evolve one tiny mutation at a time, and so the population can’t adapt when circumstances change. A diverse population offers the opportunity to try new combinations in response to a changing environment.
- Diversity offers resistance to epidemics. A parasite is a simpler organism that is usually optimized to prey upon one particular genotype. In a homogeneous community, the parasite spreads seamlessly from one individual to the next; in a diverse community, the parasite has a harder time.
I constructed two separate computer models, based on these principles and also structured in ways that made the two models as different as I could conceive them.
The results showed that, by adjusting parameters, I could make each of these models produce senescence as an adaptation, with rejuvenation tied to gene sharing.
If you’re skeptical about computer models, as I am, you’ll want to know whether the particular parameters that are friendly to selection for senescence are typical of what we find in nature. But the models are abstract and idealized, difficult to relate to complexities in the real world — let alone the real world two billion years ago, when sex first appeared. So these models are just a start, just a suggestion.
The big picture
If you’re a paramecium, you can clone yourself for awhile, but your clonal lineage is a dead end. Only the community survives long-term, and not the individual.
For multicelled creatures, we can’t clone ourselves at all. Each of us is destined to be a one-off experiment in combinatorics, never to be repeated. It is the gene pool that has Darwinian staying power, not the particular combination that is you or me.
This principle has deep consequences for the whole process of evolution. It means that, in the long run, it is the fitness of the community that matters. Individual fitness is fated to be a transitory phenomenon. It is community fitness that can persist, can compete, can flourish in Darwin’s competition.
This is the opposite of the “selfish gene”. Richard Dawkins (and R. A. Fisher 46 years before him) had it exactly backwards. According to the selfish gene dogma, which dominated evolutionary theory from 1970 onward, Darwinian competition takes place primarily at the individual level. That was the old theory. But sex changes this. The meaning of sex is precisely to lift the level of evolutionary selection from the individual to the community.
Life is evolved for cooperation, just as much as for competition.
Discover more from Josh Mitteldorf
Subscribe to get the latest posts sent to your email.
Well a random mutation or mutations in the ability of individual paramecium to replace their macronucleus from the archived micronucleus, on the face of it a bad thing, ended up allowing those mutants to have more surviving offspring. I don’t think there is a particularly complex mystery to that. You could argue that all ageing evolved that way, whether the mechanism is telomeres, DNA errors, epigenetic regulation, whatever. It is more interesting to think about how nature seems to always favour co-operation on greater and greater scales. Even zooming out into the human realm, selfish individuals may be able to have more children in the short run, but it is the co-operative societies that end up winning in the long run. It is almost enough to make one believe in god.
The part of the mystery I didn’t convey very well is the sense in which the Fisher paradigm is correct. I spent 5 years (1996-2001) modeling different schemes for evolving senescence via selfish gene mechanisms, and this convinced me that the problem was, indeed, deep.
In 2001, was finally able to model senescence emerging from an evolutionary model in the context of an ecology where a breeding community shared a common food source. This breakthrough influenced my thinking from that day to this. I learned the ways in which the Fisher model is an accurate representation of reality, and other ways in which it is not.
In my view, there is still much unexplained about how the diversity of life has been created, and how the Darwinian model must be expanded to bring it into harmony with the nature we observe.
On The Other Hand – consider the planaria (a flatworm). It can have sex, but it doesn’t have to. You can cut up a planaria into at least 289 pieces and each piece will form a new planaria. They seem to be immortal
You’re right that planaria seem to be a special case, and hard to understand. Michael Levin talks about how messed up their genomes are, after thousands of generations of mutating and reproducing from pieces of themselves, and how strange that they can produce perfectly healthy worms where different cells have different genomes and even different numbers of chromosomes.
For a more detailed look at the cell level in planaria, see the following article – https://www.cell.com/cell/fulltext/S0092-8674(18)31233-9?_returnURL=https%3A%2F%2Flinkinghub.elsevier.com%2Fretrieve%2Fpii%2FS0092867418312339%3Fshowall%3Dtrue
There seems to be stem cells of all different type spread throughout the planaria’s body. Still, from a group population dynamic, the planaria seems to be the complete opposite of protistas. Nature is a wonderful and strange place.
Super interesting Josh, I really liked this post.
“For multicelled creatures, we can’t clone ourselves at all. Each of us is destined to be a one-off experiment in combinatorics, never to be repeated. It is the gene pool that has Darwinian staying power, not the particular combination that is you or me.”
Just wanted to point partenogenesis as exception, it happens in many insects fishes and snails, several reptiles and amphibians and still a bunch of birds have been documented as well. Femnales reproducing just because of lack of mate.
It is almost disturbing.
Yes, so true…
Ben test comment