
Eating less is the best-tested and surest way to a younger body and an increased life span. But it’s a hard discipline to maintain, and many of us would welcome an easier alternative. Perhaps we can realize some of the benefits applying a more temporary exercise of willpower, with intermittent fasting. It’s counter-intuitive, but seems to be true, that health and longevity are better served by clumping up our food consumption (feast and famine) than by spreading food consumption evenly through the day and through the week. The topic is controversial, and the evidence is not just unclear, it’s contradictory. The bottom line is that it is worth trying. Some people who drift away from diets involving consistent discipline find that they can comply on an intermittent schedule. Experiment with different schedules, because individual response varies widely.
The link between restricted diet and longer life goes back at least to Benjamin Franklin: “To lengthen thy life, lessen thy meals,” said Poor Richard. Formal experiments on mice established the benefits of calorie restriction in the 1930s. But calorie restriction remained a backwater of research, unknown to most biologists, let alone the public. Beginning in 1946, intermittent fasting was introduced as an alternative lab regimen that required less measuring and monitoring. These early experiments suggested great promise for cancer prevention and life extension by restricting access to food on various schedules. This was exploratory work, and the experimenters were not keeping proper controls, or varying just one factor at a time, so it’s hard for us now to fit those results into the base of later experiments.
In the 1980s, caloric restriction was already well-established as a robust way to extend life span, and alternate-day feeding and fasting was re-discovered. In the best outcome, rats lived 83% longer when fed every other day. Looking back from today, we would want to ask: how much of that benefit came from lower consumption overall, and how much from the schedule and periods of fasting? Did they eat much less overall? Or did they gorge themselves on the in-between days and make up for lost time? Though no records were kept of the animals’ food consumption, the results seemed to be too good to be due to reduced calories alone. The schedule seemed to be at least a contributing factor in the success of the diet.
Some species are adapted to graze continuously, while others have a much greater capacity to store food and digest it slowly. Even within the same species, different varieties may react very differently to alternate-day diets. A study by Goodrick (1990) highlighted the different responses of two strains of mice. Both were put on the same every-other-day regimen. Both maintained their body weights, with no significant differences from control animals fed ad libitum. But one strain enjoyed consistent life extension, and the other suffered a slight decrease in life span from EOD feeding.
Experience with people
People have a hard time adapting to no food at all every other day, so the four regimens that have been suggested for humans are
- Concentrating food intake in a portion of each waking day, or eating one meal a day, or fasting every day for 12-16 hours
- Fasting or eating lightly on alternate days
- Fasting one day a week
- Longer fasts of several days, less frequently.
Long-term human studies of EOD fasting and longevity are not available. (People don’t like to live in cages, or eat the same thing as other subjects for years on end.) So instead we look for hints in the short-term metabolic response to intermittent fasting. The metabolic connection of diet to aging is mediated through the insulin metabolism, so it is logical to ask what are the effects on insulin levels, insulin sensitivity, and blood glucose. EOD fasting shows benefits comparable to CR for some but not all of these. Higher levels of HDL (“good cholesterol”) have been reported for humans and animals, and there is good support for lowering of cancer risk in animal studies, but no data yet for humans. In the most pessimistic study, combining calories of 3 meals into one big meal in the evening had a negative effect on the insulin metabolism.
Protecting the Aging Brain
An intriguing benefit of the EOD regimen seems to be an increase in BDNF (brain-derived neurotropic factor). This is a hormone that promotes new nerve growth in the brain, and presumably is related to the ability to learn. BDNF levels decline with age. There is good evidence that some of the benefits of EOD fasting and of CR generally are not direct effects on the metabolism, but are mediated through the nervous system. Maybe you have to feel hungry to receive the health benefits. The BDNF connection supports that idea.
Weight loss – a good indicator
For some people, losing weight is an end in itself, and a primary goal. But even if your focus is on other benefits (life extension or long-term health), weight loss is a good sign that the diet is working for you. If you are not losing weight, there still might be benefits from intermittent fasting, but it is harder to know.
For most people, trying to eat less via willpower is counterproductive. A large majority of people who stay on a diet using discipline find that they regain more weight than they lost after their resolve runs out. Such statistics, of course, always apply to other people, and each of us knows we can do better. And some of us are right.
Advice
Experiment with a chosen fasting schedule. Stick with it at least a month to give yourself time to adapt, and to average over varying life circumstances. The right diet for you is the one you can live with. When you find a schedule that is right for you, you will enjoy lightness and alertness; you won’t feel deprived or resentful; there will be satisfaction in caring for yourself well, and sensing it.
Krista Varady of Univ of Illinois has a research program helping people with every-other-day diets with 7 years of experience. Varady reports good success using a schedule of alternate day dieting, in which subjects eat one meal on the in-between days, a normal lunch of 400-600 calories depending on body size. On the eating days, she says subjects average 10% more than their usual diet, but do not pig out regularly, once they get used to the routine. Some subjects drop out of the program, but for >80% compliance is good. She says many people are able to stick with this EOD schedule long-term, to lose weight quickly and keep it off.
In summary, intermittent fasting is likely to work, but not for everyone. Only personal experimentation can tell you if you’re able to accommodate to fasting, if it can fit into the demands of your life, if you tend to overeat before and after.
Personal note: I’ve found I can live with a complete fast one day a week (usually on Thursday). I also try to extend my overnight fast at least 12 hours. I don’t think I eat less overall, because I actually gained a couple of pounds when I started doing this in 1997. But weekly fasting offers a kind of sabbath that feels right to me, makes me a little less focused, less verbal, less patient but more introspective.
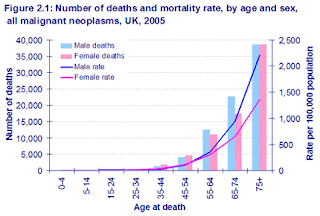 Most researchers have explained this by positing that mutations in a cancerous lineage accumulate over many years until the last safeguard is gone, and the cell can wreak its havoc. But this remains purely hypothetical, since an increase with age of “partially converted” cells has never been observed. Meanwhile, it is well known that
Most researchers have explained this by positing that mutations in a cancerous lineage accumulate over many years until the last safeguard is gone, and the cell can wreak its havoc. But this remains purely hypothetical, since an increase with age of “partially converted” cells has never been observed. Meanwhile, it is well known that 