
Of the many species that outlive their fertility, lab worms (C elegans) take the cake. They stop laying eggs after two days, but they can go on to live for two more weeks. Did they miss school the day that the teacher was lecturing on Darwinian fitness? Or perhaps do the elders of their community have a message for their grandchildren?
In the paper that launched the modern evolutionary theory of aging, George Williams [1957] predicted:
“(6) There should be little or no post-reproductive period in the normal life cycle of any species.”
Not so long ago, evolutionary scientists thought that human menopause was unique in the biosphere. A whole industry arose explaining the “grandmother effect”: Playing the game they like best, population geneticists would try to concoct circumstances under which a woman’s “selfish genes” could leave more copies of themselves by not reproducing, but instead freeing their bearer to raise her grandchildren, while their mother went out hunting and gathering.
Then a literature began to appear in which other animals were discovered to outlive their fertility. But this didn’t stop them. Seldom do facts on the ground discourage an enterprising evolutionary theorist.
In several whale species, the female undergoes menopause and continues to live for several decades thereafter [McAuliffe]. Elephants, lions, and baboons experience menopause [Packer], and all these animals might plausibly be argued to relate to their grandchildren.
But some other animals, not so much. As early as 2004, menopause was recognized as a “generalized mammalian trait”. Beyond mammals, hens and guppies both stop laying eggs. Yeast cells stop budding.
James Vaupel’s research group published a comprehensive survey of animal “life histories” [2013], in which they charted fertility and mortality over a lifetime for dozens of species of plants and animals. The blue lines map fertility, and the grey background curves show survival.
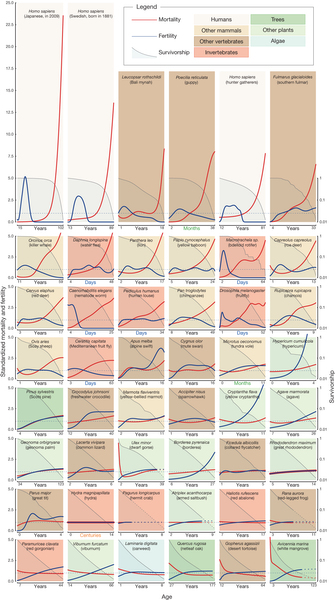
In these 48 plots, C elegans worms, the common lab roundworm, stands out as living most of its life with no fertility at all. Why would that be?
My principal contribution to the evolutionary theory of aging has been to cite demographic stability. Population homeostasis doesn’t come for free; in fact all ecosystems would blow up if their constituent species were evolved to live and reproduce as copiously as possible. In general, aging has evolved as nature’s way to prevent population overshoot, to avoid extinctions that come when populations crash, and thus to keep ecosystems stable and productive in the long term. This is my theory paper, with Charles Goodnight, on post-reproductive lifespan as a demographic adaptation.
The life cycle of C elegans in nature
C elegans worms are hermaphrodites. For the first day of their lives, they are male, and generate a few hundred sperm cells. Then they become female, and start laying eggs, one by one, fertilized by their own stored sperm. But after two days, they run out of sperm and can’t fertilize their own eggs. They become infertile, unless…
One worm in a thousand is not a hermaphrodite but a male. It lays no eggs, but produces oodles of sperm, which can fertilize the eggs of a hermaphrodite worm. But she has to be very lucky to find a male, because males are so rare. So this is an interesting footnote, but probably irrelevant to the reason worms are programmed to run out of sperm and discontinue fertility. The vast majority of hermaphrodite worms just live out their lackluster lives. Without sex, is life worth living?
When they are not growing in a Petri dish, C elegans worms live on rotting fruit in the wild. One piece of fruit on the ground can feed millions of worms over a period of several weeks. But how are they to get to the next piece of fruit?
If the worm is fat and happy, it lives 2 to 3 weeks. But if the worm is starved, it has a special adaptation. When food is scarce, it can go into a diapause stage called dauer, in which it doesn’t eat and its metabolic rate is very low. As a dauer, the worm can live for many months. The dauer can be ingested by a bird and survive digestion.
Carried by the wind or hitching a ride in a bird’s intestine, dauers are the worm’s plan for getting from one piece of fruit to the next. The “fitness” of a worm is probably better measured by its probability of finding a new piece of fruit than by the number of offspring it produces. Fitness is a property of a worm colony, not of a single worm. The fitness of a colony is related to the number of dauers that are sent out to explore the environment for the next piece of fruit.
Do the math — at several hundred worms per generation, the first three generations will do fine. They will be able to gorge themselves on fruit. But the fourth generation would be hundreds of times larger. The worms would start to grow, only to find that there are so many of them competing for the fruit that none have a chance to grow to maturity.
This is why the ability to dauer is not just useful but essential to the worms’ collective survival. Larval worms detect food scarcity, and they flip a switch into dauer mode. They cast their future to the wind, or commit themselves to the gut of a hungry bird.
There is a danger, though, that they might miss their timing. If they are starved right out of the egg, the worm is not yet large enough to form a dauer. But one day out of the egg, the worm is already committed to maturity, and it is too late to dauer. The dauering window is just a few hours long. It is easy to miss.
Here is where grandma can help. In a worm colony that is in its fourth generation, there are already hundreds of thousands of elder worms, post-reproductive but still squirming. These worms saturate the nest with dauer pheromone. The pheromone signals the hatchlings of the fourth generation that, even though it may seem that there’s plenty of food for now, the generation size is so high that the food supply is in danger of being exhausted before they can grow to maturity. The pheromone encourages the young larvae to go into dauer rather than take their chances on maturity and reproduction in present conditions. Dauer pheromone from the oldest worms is a signal both of crowding and of colony age. Exponential mathematics is what is threatening the worm population, and exponential mathematics is what potentiates the enormous increase in dauer pheromone that inform the larvae that their best prospects are to leave home and seek their fortune in the countryside.
This last paragraph is my personal theory, not yet fully confirmed. For a decade, I have been looking for someone to actually measure dauer pheromone from elder worm populations, and no one has taken up the challenge in this form. But from two labs where I have worked, there is partial confirmation. These are in the Beijing National Institute of Biological Sciences (Meng-qiu Dong and Wu Gang) and at Washington University of St Louis (Kerry Kornfeld and Andrea Scharf).
Evidence for the grandmother worm theory
NIBS Beijing
In 2017, while I was a visiting scholar at the National Institute for Biological Sciences in Beijing, my host, Meng-qiu Dong, was kind enough to design two experiments with grad students Wu Gang and Peng Lei.
First they raised infertile worms synchronized by age, so they could extract liquid from their growth medium when all the worms were a fixed age. They confirmed that dauer pheromone could be extracted from the medium, and that the quantity peaked for 5-day-old worms. Larvae grown in the medium where 5-day-old worms had lived were more likely to dauer than larvae in a control medium. 5-day-old worms are grandparents, but not yet great grandparents.
Second, they grew worms to old age on half a Petri dish, and then grew a fresh generation of worms on the same Petri dish. As the young worms moved around, they tended to avoid the half of the plate where the old worms had lived (compared to a control dish where no worms had previously grown).This suggests the worms are instinctively avoiding crowding.
Results written up informally here were never published because they are highly preliminary. I think the hypothesis warrants further experimentation.
Wash U St Louis
In the WashU lab of Kerry Kornfeld, my job was to model the worm’s physiology and life cycle based on individual measurements of their growth, food consumption, maturation, dauering, egg-laying, and death. My computer model aggregates the worms’ individual behavior to predict population growth and population crash in a test tube where food (bacteria) is added on any given schedule. In our first published paper, we reported good agreement between the model and population curves as measured with worm counting software.
But there was one persistent disparity between the predicted population curve and the lab results. With daily feeding, the computer model predicts that population should reach a peak on the 10th day, before declining precipitously and then leveling off with smaller oscillations. But in the lab, the worm population peaked on the 13th day, a lower peak followed by a less calamitous decline.
Secretly, I was delighted. We had been working on this project since 2014, and I had predicted at the beginning that our most significant discovery would be finding that we could not accurately model the worm population without including pheromone communication in the model. I created and refined a computer model for the WashU team that had only individual worm behaviors. But on the side, for my own curiosity, I was adding a parameter that controlled worm communication mediated via pheromones.
I tested the bare model (no pheromones) by varying parameters within the experimental range and beyond, but the time to the first population peak was very insensitive to all my parameters. This is because the population is increasing exponentially, so between the 10th and the 13th day, the population explodes by almost 100x.
But one change in the model that is capable of reproducing lab results is to increase the probability of dauering as the population matures. This is indirect but tantalizing evidence that the old worms really are sending a chemical signal that warns the young larvae not to try to grow to maturity, but to dauer instead.
My theory and why it is significant
Chemical signals from the grandmother worms alert a late generation of larvae that they should not try to grow up, but go into suspended animation. This softens the incipient population crash and assures a bumper harvest of dauers that spread through the neighborhood in search of a new home.
My hypothesis is that populations are evolved for homeostasis, to avoid population overshoot and crashes that can lead to extinction. Roundworms in nature are subject to highly variable food availability, and they are highly adapted to protect against population crashes.
The “selfish gene” model is still the prevailing paradigm in academic evolution. It is difficult to understand post-reproductive lifespan within this paradigm. If confirmed, this would be an example of an adaptation at the population level in which individuals cooperate to guard against extinction of the community. There is an established communication system, even in this primitive animal, explicitly for this collective purpose.
Discover more from Josh Mitteldorf
Subscribe to get the latest posts sent to your email.
This is a special case and, therefore, cannot serve as evidence for your theory. You are making the same mistake as the neo-Darwinists who select a couple of examples that appear to support their theory and ignore the overwhelming counter-evidence.
The most fundamental principle of good science is this: Instead of trying to verify your theory to confirm it, focus all your effort on falsifying your theory. Quantum physics is one of the most well-confirmed theories in science’s history not simply because it has been useful in explaining a couple of phenomena, but because countless efforts by scientific giants, such as Albert Einstein, failed to falsify it.
I guess the main question here from your point of view Josh, is are the worms releasing these pheromones in a selfless manner to help wormkind, or is it just a side effect of a lot of old worms crowded together triggering the young to go to sleep/look elsewhere?
On a side note, I think the selfish gene vs selfless group selection arguments are not helpful terms. Humans are nerve-based creatures operating on a (psychological) level above genes, and so are of course capable of being extremely selfless, and through this, creating extremely successful societies. Worms certainly aren’t able to do this, and hence any selection method that uses selfish or selfless terms is unhelpful as it draws IMO an invalid parallel with humans. We can of course discuss the human condition, but that is quite different to the worm condition.
It’s clearly an altruistic adaptation, but not hard to understand since the whole colony is inbred so the genes that are being promoted are everyone’s genes. It’s altruistic because (1) there’s no benefit in stopping my own reproduction as I get older, and (2) the limited study we have shows that there is a peak in pheromone production when the worms are a few days past menopause — past the time when they can benefit directly. But I emphasize that that the worms who benefit are closely related offspring and cousins in a colony with low genetic diversity. “Fitness” makes most sense as an attribute of the colony as a whole, and it’s measured by the ability to seed a new colony.
You say there is no benefit in stopping reproduction, but I’d look it the other way around and say, why don’t post menopausal worms just die? It must be more beneficial to hang around signalling starvation than to die earlier and free up resources; there must be a benefit in spreading to new colonies rather than enduring in the current one. At this stage I think it is more useful to consider the worm colony as one big organism rather than individuals. Otherwise the tendency is to compare individual worms to individual humans, which then infects our thinking with ideas of selfishness or selflessness as I’ve already discussed.
I’d say that there are in between states of the unit of selection. This is DSWilson’s signature idea of Multilevel Selection (MLS). In other words maybe the bee colony is a primary unit of selection, and maybe the individual amoeb is a primary unit of selection, but even in bees there are individual effects and even for amoebae there are social effects.
Humans are, of course, the case we care most about, and we are also the most complicated case. There is individual selection and there is communal selection for humans and they take place simultaneously, sometimes in the same direction, sometimes fighting against each other. Then there are megalomaniacs like Genghis Khan and Atilla the Hun and Bill Gates who end up pushing group selection in directions that are not beneficial for the group.
I think the likes of you and Wilson are treading a very fine line trying to explain human behaviour in terms of genetics. It is like trying to explain chemistry with quantum physics: overly complicated and one level too deep in the weeds. It makes me think that maybe this is the wrong approach for understanding ageing, too.
This article is about worm behavior, not humans.
And quantum mechanics provided an understanding of the periodic table and of chemical bonds that we never had before Pauli, Born, Lewis, and Pauling.