
“If aging is driven by damage, then damage must accelerate aging. If hormesis induces damage and slows down aging, then aging is not driven by damage. So a straightforward explanation is that aging is not caused by accumulation of molecular damage.” —Mikhail Blagosklonny
Last week, I wrote about hormesis*, which is the strange tendency of living things to languish when conditions are ideal, and thrive best when life is tougher. Lab animals are found to live longer when they are underfed, exposed to toxins or radiation or pathogens or temperatures that are uncomfortably high or low. I will follow up in this column with more about Caloric Restriction, which is the clearest example of hormesis. Then I mean to expand and clarify the statement about the implications of hormesis for evolution, and for our understanding of aging.
Hormesis is where my study of aging began, though I did not yet know the word. In 1996, I learned for the first time that many animals live longer when they are underfed, and their life span continues to increase with decreasing calorie intake, right up to the treshold of starvation. I drew the logical conclusion: It does not make sense that animals would be able to do something when they are starving that they are unable to do when they have enough to eat. If animals have shortened life spans when they have all the food they need, it means that their genes are not evolved for maximum life span. Aging must be“voluntary”, in the sense of a genetic program, crafted by natural selection. Quite simply: if animal metabolisms were “trying” to live as long as possible, then certainly “as long as possible” would be shorter when they have plenty of food, and when they don’t have to fight off various diseaes or repair damage from radiation or injuries, or expend energy in running.
This is the realization that convinced me I had a mission and a message in the field of evolutionary biology of aging. 18 years ago, it was a guess based on a single piece of information, but much that I have learned in intensive study since that time has confirmed the proposition that aging is programmed into our genes. There are many other forms of hormesis, sampled in last week’s column. Additional proof comes from the fact that the genetic basis for aging is very old. Genes that regulate life span are closely related in species as diverse as worms and mice and even yeast cells, though they represent branches of an evolutionary tree that diverged more than half a billion years ago. Of course, there are many genes that we share with these early life forms, but all of them are involved in core life processes of the cell, like replication and transcription and energy production. We can only conclude that nature has treated aging like a core life process.
And as more is learned about the physiological mechanisms of aging, it appears that some are quite avoidable, and others look like deliberate self-destruction. For example, late in life the genes that code for protective enzymes such as ubiquinone and glutathione are dialed down, while signals that spark inflammation are dialed up to the point where inflammation becomes a major risk factor for cancer, heart disease, and Alzheimer’s
Caloric Restriction Experiments
The connection between less food and longer life predates modern scientific study. Hippocrates hints at it. In 15th Century Venice, Luigi Cornaro wrote a volume titled Discorsi della vita sobria (Discourses On the Temperate Life) about his personal experiments with caloric restriction, supplemented by half a litre of wine daily. Cornaro lived to 102. Benjamin Franklin wrote in Poor Richard’s Almanac, “To lengthen thy life, lessen thy meals.” (1733).
In the depression of the 1930s, the issue of widespread malnutrition was discussed in America. How would it affect people’s health and longevity if they did not have enough to eat? Clive McCay was a young researcher at Cornell when he received a grant to study the relationship between delayed growth and life span in rats.
McCay wasn’t thinking in terms of hormesis, and evolution was far from his thought process. In fact, his framework was rooted in the (discredited) rate of living theory. He thought that perhaps growth, development and aging are all synchronized on a common clock, so that if he could delay growth and maturation, then perhaps aging would be delayed as well. In fact, the way that McCay controlled his rats’ diet was to give them just enough to eat to keep them from dying, without allowing them to gain weight. His experiments with underfeeding produced dramatic results, but they were not recognized as being important, and there was little follow-up for almost half a century.
The modern re-discovery of Caloric Restriction (CR) came when Roy Walford served as “house doc” in the Biosphere II experiment in Arizona, 1991-93.

This was to be a hermetically-sealed environment in which a team of bionauts grew all their own food, recycled all their water and even used photosynthesis to supply their oxygen. But farm productivity was way below expectations, so the crew didn’t have enough to eat. Walford noticed that underfeeding the bionauts had dramatic health benefits, though it made them irrascible.
Over the years, experiments have been done with yeast cells, worms, fruit flies and other inspect species, arachnids, crustaceans, fish, various rodents, dogs, horses, and rhesus monkeys [Ref]. A project at Washington University has followed the health histories of people who practice CR [Ref], though mortality comparisons may not be available for a long while. Shorter-lived animals tend to show greater proportional life extension, but health and longevity benefits were discerned even in the monkeys, with a life span of 25 years [Ref1, Ref2].
McCay’s guess about a biological clock and delay of development turned out, at best, to be only partially correct. CR works even if begun in adult animals. Life extension is not as great as when begun earlier, but qualitatively the effect is the same.
CR increases both mean and maximum lifespan. It’s much easier to increase average life expectancy by preventing early death than by truly delaying aging. The signature of slowed aging is often taken to be the impact on the last survivors, because that is where it can be observed most sensitively. Sometimes the difference is illustrated with survival curves that look like these two:
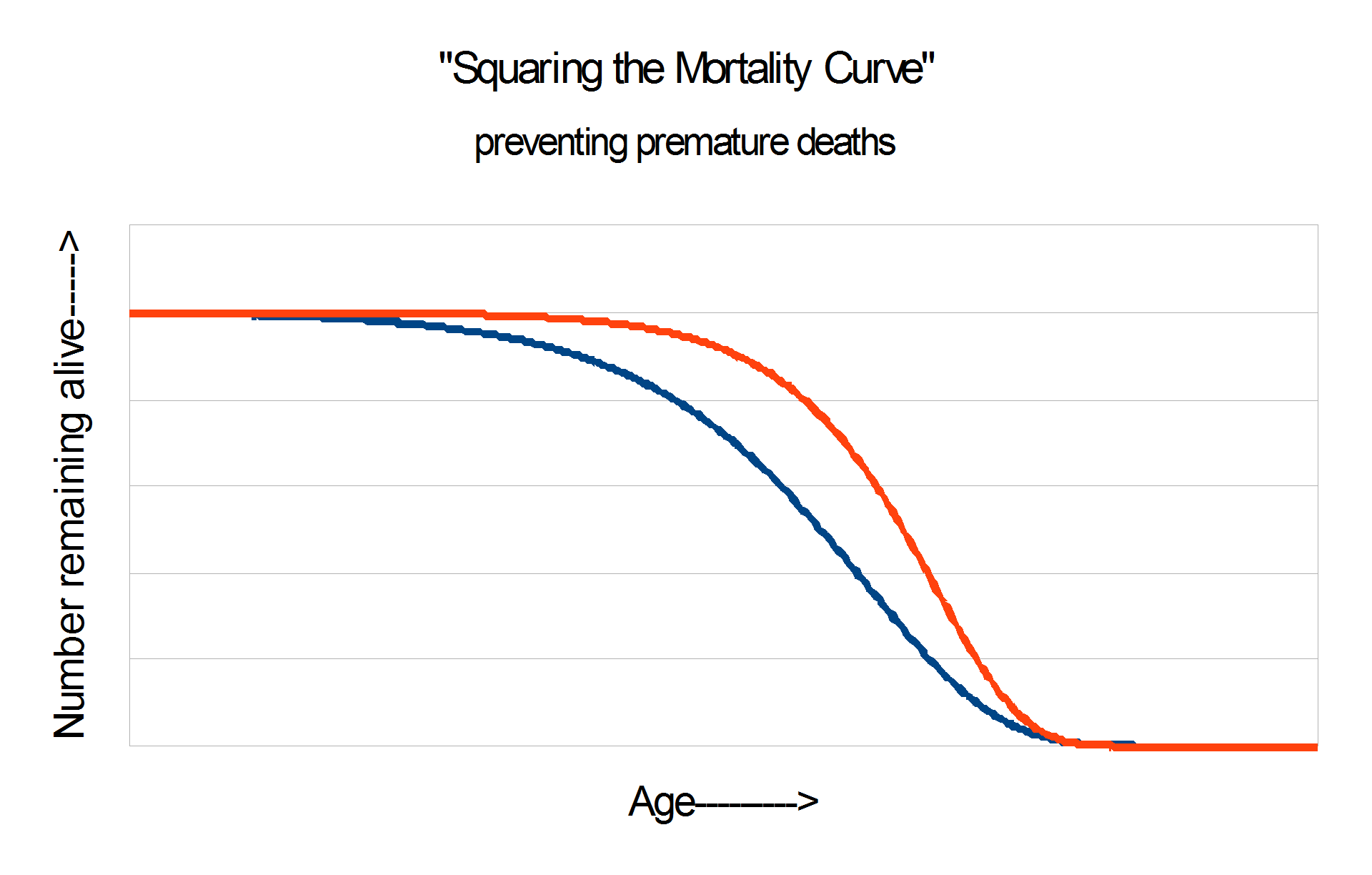
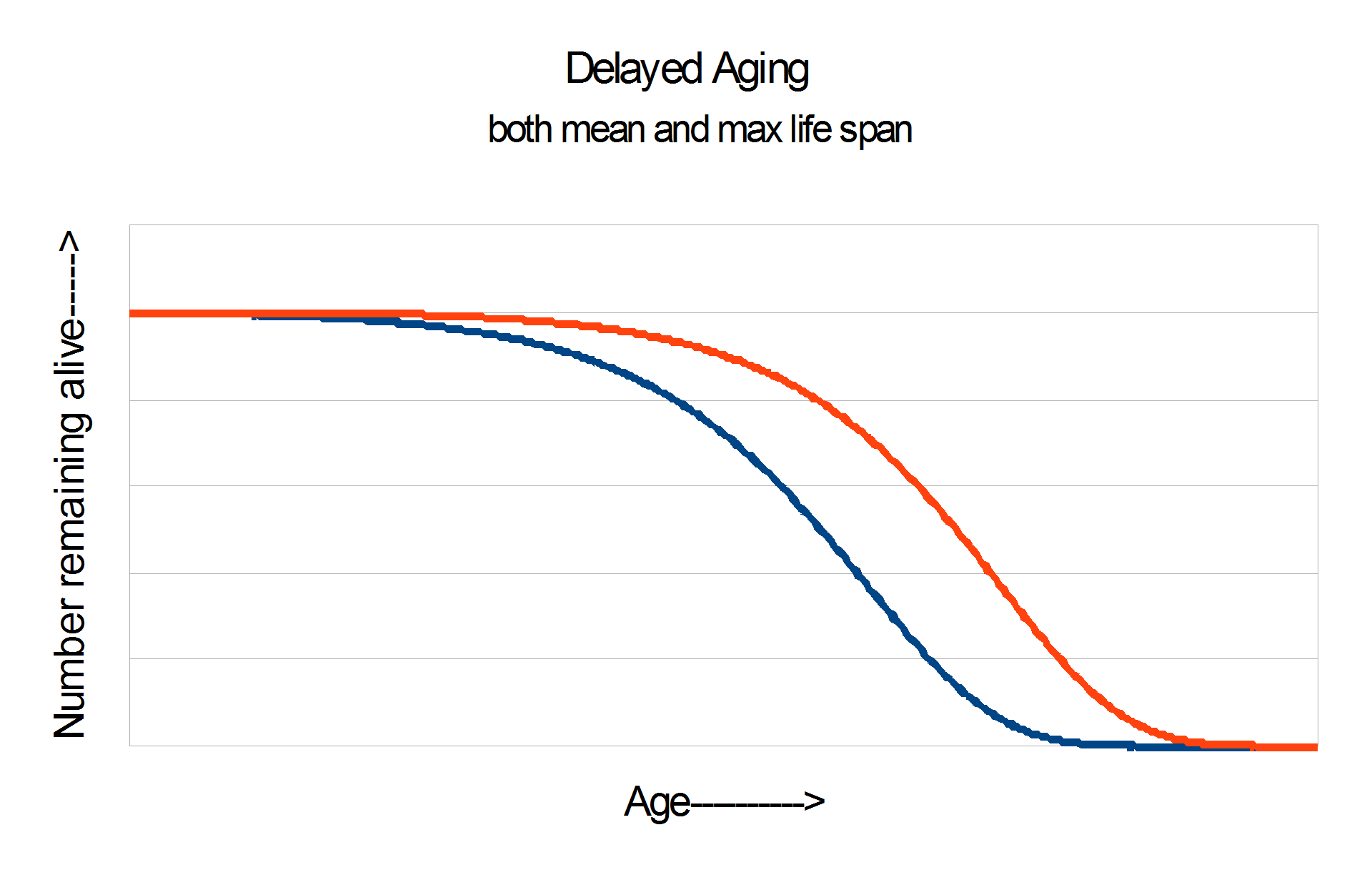
Results of caloric restriction experiments typically look like the graph on the right, which is regarded as the “real McCay”.
Short-lived and simpler animals tend to show (proportionately) a better response to CR than long-lived animals. Lab worms fully-fed live only 10 days, but if starved early in life, they go into a suspended state called dauer, which is something between hybernation and a spore. Dauers are extra tough and resistant to heat, cold, dehydration, and other things that normally would kill a worm, and they can survive up to four months without food. A dauer is alive just enough to detect food and water in its environment, and when it does, it picks up life and growth from where it left off
Lab mice normally live two years, and with a severely restricted calorie intake they can survive for three. Dogs typically live an extra 2 years on CR. The longer the life span, the less the proportional gain. Some reporting in the popular press would lead you to think that the long-term experiments in Rhesus monkeys found little or no gain in longevity, but I’ve viewed the results as positive, though there were plenty of interesting complications.
CR in humans has attracted many enthusiasts. It’s a very individual practice. Some people have been overweight most of their lives, and are trying to get their weight under control; others are very skinny, and disciplining themselves to be yet thinner. There are major differences between people who take lots of supplements and people who go “natural” or “paleo”. The greatest diversity is around exercise: Some people on CR are also doing high-intensity exercise programs. Exercise independently increases life expectancy, and has health benefits in the present. Exercise is the most robust and long-lasting anti-depressant known to psychological science. (Personally, I tend to swim with this school.) There are others who avoid exercise, because exercise burns calories, and this inevitably leads to more eating. They are tracking calories rather than weight. People in this school often base their thinking on the fact that exercise increases only mean life expectancy but not maximum life expectancy (see above two graphs), so it does “not truly slow aging”. My response to this is to re-phrase the issue: Yes, there are a lucky 2% of people who will live a long time whether they exercise or not, but there are 50% of people who delude themselves into thinking that they are in that 2%. Read stories and get a sense of the culture at CRSociety.org.
Of course, there are many other people who have decided that the discomfort and discipline of either calorie restriction or exercise are too high a price for the (substantial) improvement in health or the (modest) increase in life expectancy.
An important implication of the CR experiments is that life span is extended not through some physiological action, but through the effect on signaling. This carries a powerful message for evolutionary understanding. Aging is not simple accumulation of damage, because the damage can be delayed by mere chemical signals, or instructions to the living metabolism. Here’s a sample of some of the studies that demonstrate this.
- From Cynthia Kenyon’s UCSF lab, it was reported that merely sensing (“smelling”) food without actually ingesting the calories was sufficient to cancel some of the life extension from food restriction in worms.
- The same lab discovered one of the genes (DAF16) that served as point man for the CR response. They were able to genetically engineer worms that had the DAF16 gene only in their muscle cells, or only in their nerve cells, or only in their digestive cells. From experiments with these mosaic worms, they were able to show that it was the nervous system that dictates the life span.
- In experiments with rodents, every-other-day feeding extends life span, even though the animals eat so much on the in-between days that they get almost the same number of calories as fully-fed animals.
Theoretical Understanding of CR – what they got right, and what they missed
The first (and now standard) explanation to relate the CR adaptation to evolution was by veteran mouse geneticists at Jackson Labs in Bar Habor, where different genetic strains of lab mice are bred for labs the world over. (This was 1989! For the first half century in the history of CR research, no one had thought to connect this major adaptive response to evolution.)
Harrison and Archer theorized that the CR response originated as an adaptation to famine. During a famine, it would not pay an animal to reproduce, because its offspring would probably starve. Better to conserve resources and try to live out the famine. In fact, those animals that managed to survive past the famine would have an opportunity to deliver their offspring into a world where food was once more plentiful and competition for that food was thinned by starvation. Their genes would seed a renewed population with enhanced opportunity for success.
In my view, this is exactly right. But they did not take the next step to explore the crucial (and controversial) question: What enables animals that are starved to live longer? This vague idea of “conserving resources by not reproducing” helps explain why females don’t reproduce when they’re starved, but perhaps that doesn’t need much explaining. The point is that
- males still do have most of their fertility when they are calorically restricted, and
- females that don’t reproduce don’t live longer. (They may live shorter.)
This shows that the life extension is not caused by restraining reproduction. There are two independent signals, one that shuts off fertility and one that turns on longevity. Here’s my paper on the subject.
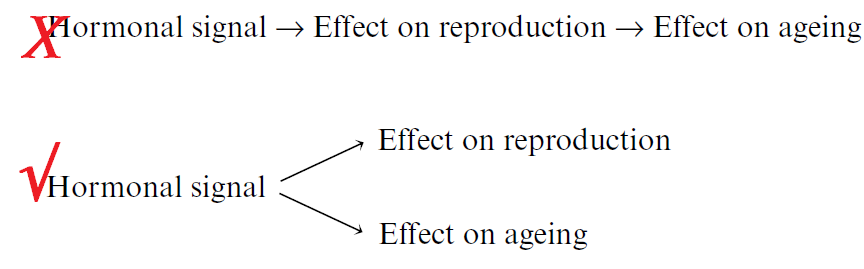
Once again, the message is that aging is controlled by signaling, and the signaling for aging rate is separable from the signals that control reproduction. The idea of “conserving resources” is a red herring, because aging is not caused by scarcity of resources.
Why have so many smart biologists missed this message?
Most biologists have been led astray by evolutionary theory. This happens both directly (because they understand the deep conflict between the ideas of evolved aging and the Selfish Gene) or indirectly (because they are immersed in a cultue that reveres nature, and they believe nature has made each individual just as strong and as durable as possible).
Darwin’s vision of natural selection says that the name of the game is to prevail in evolutionary competition, to survive and to leave more offspring than your compatriots. Aging destroys your competitiveness and cuts off your fertility. Aging is the opposite of fitness, and the idea that aging could be “adaptive” is absurd on its face, a non-starter.
But that’s what nature is telling us, so we’d better find a way to re-fashion our theories to accommodate reality.
As I mentioned last week, Harrison and Archer’s theory about CR actually points us to the place where evolutionary theory needs to be corrected. Animals are not just adapted to be prime individual competitors, but also to be members of a stable ecosystem. Ecosystem collapse is a very real danger. It can happen in a single generation, and presumably that is exactly what has happened many, many times in the deep past. As a result of natural selection among ecosystems, we have today a world of relative stability and homeostasis. I have explained in a previous columns my Demographic Theory of Aging, and I have academic articles on the subject [2008] and [2012].
The bottom line is that starvation in nature is ubiquitous, and it is especially dangerous to the community. Starvation tends to happen to happen to everyone or no one. (If I can’t find anything to eat, then chances are that you can’t either.) Natural selection has responded by designing individuals to die on an individual schedule, so they don’t all die at once. Even better, natural selection has arranged to relax the schedule when starvation is afoot, because at times when many are dying of starvation, the last thing the community needs is for more to die of old age.
* I am grateful to readers who pointed me to two factual errors that made such good stories.
- Paraquat is not the same as Agent Orange, which became infamous in the Vietnamese war. Both are defoliant poisons, and strong oxidants, but they are chemically quite different.
- Survivors of Hiroshima and Nagasaki did not have lower cancer rates later in life. They had elevated cancer rates, as you would expect. Some epidemiologists claim to see a signature of hormesis in the data of those whose radiation exposure was quite small.
…thanks for keep me honest. I think the thrust of the narrative is intact.
Discover more from Josh Mitteldorf
Subscribe to get the latest posts sent to your email.
Excellent article Josh. I agree with just about everything you say, except your first quote from Blagosklonny suggesting that hormesis is damage and therefore if it counter-acts aging, then aging cannot be damage. As you know, I certainly don’t think that damage is the ultimate cause of aging, but I do think it is a proximate cause.
As you yourself mention, the body turns down repair and maintenance systems during aging – exactly what not to do if you want to continue life. Hormesis, I believe, by introducing repairable damage (reparable); and it’s my belief that induces the repair and maintenance systems that have otherwise been down-regulated during aging. There is enough capacity to fix the damages that initiated the process and more left to correct defects that were already there before the hormetic treatments began, resulting in a net repair of damage and a turning back of the damage accumulation cellular clock.
Hi Josh. The quote by Mikhail Blagosklonny is illogical and not correct. It is a fact that aging (degenerative harmful dysfunction) generally accelerates with the passage of time. Hormesis induces damage and slows down some aspects of aging but this damage is seen more as a mild ‘challenge’ and not as harmful destruction. Hormesis is based on a ‘low-dose stimulation, high-dose inhibition’ principle. This is a crucial difference between the two meanings of the term ‘damage’.
Of course, hormesis depends on the damage being at a low level. But I think Blagosklonny is saying that the hormesis phenomena demonstrate that the body is not doing its best to resist damage during “normal aging”.
Natural selection works on individuals not groups. Individual adaptation.
Yes, sir.
In 1871, I wrote
“A tribe including many members who, from possessing in a high degree the spirit of patriotism, fidelity, obedience, courage, and sympathy, were always ready to aid one another, and to sacrifice themselves for the common good, would be victorious over most other tribes; and this would be natural selection.”
“In 1871, I wrote”
Oh, Josh. I was already agreeing with you before you revealed your true age… =)
Selection works on genes, not individuals.
Why It’s Time to Lay the Selfish Gene to Rest
Essay by David Dobbs
Group selection?
Unconvincing; evolution doesn’t work this way. See, for example:
http://edge.org/conversation/the-false-allure-of-group-selection
The set of genes that replicates best cares about its own replication, not that of the tribe aka some distantly related sets of genes. These are two very different objectives.
Yes – what you articulate is the common wisdom of the great majority of evolutionary biologists. The dominance of individual selection is such a deeply established system I won’t attempt to rebut it in this space. Instead I’ll refer you to Unto Others by David Sloan Wilson, and, if you are mathematically inclined, to Michael Gilpin’s monograph, Group Selection in Predator-Prey Communities.
If I may indulge in a one-line hit-and-run, I’d ask you how you propose to account for the fact that most higher animals reproduce sexually, giving up fully half their individual fitness, when they might have the two-fold advantage of hermaphrodites or parthenogenecists.
I notice your books are written in 1975 and 1999. That’s beause the controversy over ‘group selection’ is settled nowadays.
You can find some biologists that still support ‘group selection’, but the empiric data/numbers don’t agree, and, as such, are called out on it:
http://whyevolutionistrue.wordpress.com/2012/03/08/new-yorker-article-on-the-kin-selection-flap/
Of course, that doesn’t stop said biologists from being butthurt – but thay’re always light on arguments:
http://www.rationaloptimist.com/blog/ants,-altruism-and-self-sacrifice.aspx
Of course, in the link I posted above, I – well, Steven Pinker – provided a synthesis of how ‘group selection’ is debunked from the POVs of biology, evolutionary psychology – see his references if you want to run the numbers.
Yes – there is no shortage of big names like Steven Pinker and Richard Dawkins who still dismiss group selection, but there are now some big names who actively promote group selection. E. O. Wilson and Martin Nowak are the latest to turn around.
In the end, scientific truth is not determined by a vote of big names, but by logic. There are many widespread phenomena in biology that can only be explained by their group benefit: sex, a genome structured for evolvability, the maintenance of diversity, ecosystem stability…and aging.
Although you are discussing individual and group selection, it is at the level of an organism’s genome.
I’m not an evolutionary biologist, so I could possibly be missing some crucial detail, but all that I’ve read only suggests that the genome is a “dominant”(acknowledging the possibility of epigenetics, cultue, and stuff) unit of replication. Thus, a genome is only asserted to be a non-strict superset of the smallest unit(s) of replication. I have never read anything about whether the genome is the smallest unit, or whether it is a superset.
If the unit of replication is only a subset of the genome, and when reproducing sexually the unit of replication reproduces, but not the entire genome, then individual selection, but not at the level of the entire genome, would agree with sexual reproduction.
* I’ve read about group selection, and I think they’re very plausible for sexual reproduction.
Correction:
“and when reproducing sexually the unit of replication reproduces, but not the entire genome,”
=>
“and when reproducing sexually the unit of replication is shared/similar in both organisms, while the differences in the organisms are mostly outside the unit of replication,”
There seems here, to be a misconception regarding the difference between maturing and aging. Adolescent organisms are maturing to reproductive state and somewhere beyond that – a non-fixed period of time – enter into senescence, where a multitude of factors cascade downhill. CR has theoretically become protective during periods of starvation DURING THE ADOLESCENT AND REPRODUCTIVE YEARS only, via natural selection. Having entered the senescent stages of life however, there would reasonably be some residual protection afforded by CR, but natural selection would no longer apply (or not often – as only some “older” organisms are still able to reproduce) for adaptation to improve/reduce the effects of aging, beyond the active reproductive period.
“Dogs typically live an extra 2 years on CR”
That linked research was on Labradors which live about 11years, and it was at 25% restriction. A 50% restriction if there was proportional increase as seen in shorter lived animals, would provide 5.5years extra. 25% should show 2.75years, but there are factors that could be behind seeing only 2 years instead of 2.75years, such as maybe small sample size, in other long lived animals protein intake affects igf1 which may also be the case in dogs and could in theory also affect results if protein intake is not properly controlled.
“Natural selection works on individuals not groups. Individual adaptation.”-William. William
IT most certainly works on groups, at least groups of cells, those that form multicellular organisms. The ancestors of these cells where once individual single cell organisms in and of themselves.
“The set of genes that replicates best cares about its own replication, not that of the tribe aka some distantly related sets of genes. These are two very different objectives.”-Edit_XYZ
They’re not just distantly related, intraspecies genes are the genes with which a set of genes can couple with, outside the species there is no viable reproduction. Already, as mentioned, the sexual reproduction trait compromises reproductive capability of the individual for the ability to provide genetic diversity, which most aids the group.
Martin Blaser and Glenn Webb published a paper a couple weeks ago about the microbiome and group selection. They hypothesize that when we pass reproductive age our microbial population shifts from being protective to trying to kill us. Animals coevolved with microbial communities, and the ecosystems that selected for stable healthy host animal populations are the ecosystems that survived.
“This hypothesis may be especially relevant to isolated prehistoric human communities, where the fates of the indigenous microbes and their hosts are rigidly tethered. Host community demise would lead to microbial extinctions. As such, local communities that have equilibria between colonizing microbial populations and their hosts that maximize overall host fitness have the greatest probability of long-term survival; we have previously postulated that these relationships take the form of a Nash equilibrium.”
Their mathematical modeling found that increasing youth susceptibility to infections resulted in population crashes, but if the old people’s microbes got the best of them the overall population remained stable and healthy.
“An intriguing possibility is that the microbiome—either through alteration or not—affects host immunological function and programs its senescence or that the events are linked in a spiral of alteration, diminished immunity, more alteration, and so forth. An alternative mechanism is that chronic inflammation induced by the microbiota at particular locations (e.g., stomach, lung, colon, skin) may lead to tissue destruction (e.g., atrophic gastritis), which leads to malignancy and demise.”
http://mbio.asm.org/content/5/6/e02262-14.full
It’s an interesting possibility. We know so little about the microbiome. Does it have an independent evolutionary interest, apart from its host? Is there some kind of long-term selection operating on the microbiome, as this paper would imply?
You are reference #5 in Blaser and Webb’s paper. 🙂
If one believes in the free radical/mitochondria theory of aging; it is interesting to note that under starvation, the cell digests damaged mitochondria and recycles them as food.
Do you know something about what kind of effect long term sexual abstinence may have in organism?
Is it possible that it works as caloric restriction, some kind of hormesis, for evolutionary reasons?
If food shortage occurs in the natural environment, the chance of reproduction is reduced, so the effects produced by caloric restriction may be a way to improve the reproductive success by maintaining the individual healthy for more time as a way to improve the chance of adequate reproduction further.
So for analogous reasoning, long term sexual abstinence occurs in the nature when few partners are present, and like in the caloric restriction, it is possible that there is a mechanism to maintain the individuals healthy and increase the longevity, giving more time to try reproduction.
Is this correct? I really would like to know you opinion.
I’m pretty sure there is no data either supporting or refuting this hypothesis. It is true that many sages and ascetics report long life, and that they tend to abstinence; but you cannot conclude that brahmacharya contributes to longevity because the other ascetic practices are all tangled together.
The few studies that have been done indicate that, for animals in captivity, there is a small positive association between reproduction and longevity; and for humans, there is a more robust association between more sexual activity and lower mortality.