
Roscoff is a picture-perfect coastal town in Brittany. I have just returned from the first Monod Conference on the Comparative Biology of Aging.

Looking toward shore from the quay. My hotel is in the middle, just right of the cathedral.
The conference was opened by a theoretical lecture by Tom Kirkwood, father of the popular disposable soma theory of aging. He admonished us that evolution is a mathematical science that yields specific and quantitative information about what aging can and cannot be. These provide a powerful mathematical underpinning for the understanding of aging.
The next morning, Annette Baudisch told us that in reality, nature has produced every combination of aging strategy that you can imagine, and some that you probably never imagined. The kind of aging that humans know is gradual and accelerating, leading to death on a timetable that is predictable within about 10-15%. But this brand of aging is a small minority in nature. There are salmon and octopuses and annual plants that reproduce in a burst and then die suddenly. There are beetles and jellyfish that are able to “age backward”, reverting to a larval state under stress, then beginning life again with a fresh start. Baudisch coined the term “negative senescence” for a phenomenon that is not the same thing as this: most trees and some turtles and lobsters just grow ever larger and more fertile over decades or even centuries. There are giant lobsters that grow to 40 pounds, and there are clams you can hold in the palm of your hand that have over 500 annual growth rings. Each of these animals and plants grows progressively less likely to die with each passing year, year after year–hence “negative senescence”.
Charming and perfectly diplomatic, Baudisch overtly praises Kirkwood and the contributions he has made to the evolutionary science of aging; but in truth, she has produced more counter-examples to Kirkwood’s pronouncements than all of us combined.
I was less diplomatic, and in my presentation, I ranted about the many blatant contridictions to Kirkwood’s “precise, mathematical theory”, and in big red Powerpoint letters counseled the assembled scientists, “Don’t let the mathematicians tell you how to interpret your data.” The mathematical theory for evolution of aging is based on an early 20th Century paradigm of R. A. Fisher, in which gene frequency changes gradually while the population level and the ecology remain ever stable. We now know that ecologies change hand-in-hand with gene frequencies, on the same time scale. Furthermore, there are a dozen mechanisms of evolution that were unknown to Fisher, of which the simplistic equations of classical evolutionary theory takes no account:
- Ecological interactions
- Horizontal gene transfer
- Epigenetic inheritance
- Population cycles
- Weather cycles
- Evolvability
- Social interactions
- Learned behaviors
- Phenotypic plasticity
- Assortive mating
- Famines
- Epidemics
In every other subdiscipline of the bio-sciences, experiment is king, and theory is kept in its place. This, of course, is exactly the way science should be, and especially biology, which is so complicated that theory has only a limited role. But somehow evolutionists have carved out an exception for themselves, and when they make mathematical pronouncements that manifestly have nothing to do with the natural world, they are nevertheless taken seriously.
(Part of the problem is the illusion created by experiments in laboratory evolution. Here the theory works beautifully. But only in predicting outcomes of breeding, where the experimenter dictates the definition of “fitness”. We have no way of measuring “fitness” in nature, and have every reason to believe that it is essentially complicated, multifaceted, and completely dependent on ecological context. I introduced an aphorism that I hope will catch on: “Nothing in evolution makes sense except in the light of ecology.”*)
* Here I am echoing a great evolutionary thinker of the mid-20th Century, who famously wrote that “Nothing in biology makes sense except in the light of evolution.”
Worms with Hot Flashes
Researchers in the worm aging laboratory of Meng-Qiu Dong labeled an antioxidant protein with green fluorescent die, and discovered serendipdously that the worms have spots of activity that flash with frequency every few seconds, that you can visualize through a microscope, peaking around day 3 of the worms’ 20-day life span. Investigation revealed that the mitochondria are producing the flashes, so they’ve been dubbed “mitoflashes”. Remarkably, the frequency of flashes is correlated with the worms’ date with death two weeks down the road. Across many different strains, genetic varieties and environmental conditions, the rate of flashes at peak predicts how long the worms are going to live. Dong had the vision and insight to realize that this implies a longevity plan for the worms that is already in place quite early. The mitochondria know in advance what the life span is going to be [news article in Nature]. This supports both the perspective of programmed aging, and also the theory that mitochondria act as executioners.

False color picture of young worm, showing mitochondrial hot spots.
Sex and the Single Rotifer
Standard evolutionary theories of aging tells us that reproduction and longevity are on a see-saw, so that whenever one goes up, the other must go down. I don’t believe this, and for years I’ve been collecting exceptions. My favorite is David Reznick’s guppies. From the river pools of Trinidad, he identified two varieties of the same species: one with high fertility and long life span, the other with low fertility and short life span. It turns out that life span is determined not by individual competition to make as many offspring as fast as possible, but rather by adaptation to the local ecology. Guppies are the little kids on the block, and where there are prerdators present, their death rate can be so high that selective pressures drive them to mature more quickly, swim faster, lay more eggs, and also age more slowly. Where there are no predators, they can’t afford to be so prolific. There isn’t enough food in the small pools to finance a population explosion, and overcrowding risks the spread of fungal and bacterial epidemics
This, of course, is group selection of a kind that mainstream evolutionary theorists still deny, as they have since 1966. But in recent years, some prominent evolutionists [ref, ref, ref] have defected from the orthodoxy, and have caused a stir with the announcement of what every high school biologist knows in his gut: that cooperation and competition both have a role to play in evolutionary dynamics, and much of what we see in the biosphere is the result of a tug of war between what is good for the individual and what is good for the community.
In Roscoff, I was privileged to hear Heike Guber, a talented young experimentalist from Max Planck Institute, describe her experiments with rotifers raised in tanks. Rotifers eat algae and protozoans which they filter from the water. In different tanks, she supplied various concentrations of food, then followed them through generations to see how they evolved. The ones with lots of food evolved long life spans and high fertility; those with the slimmer diet evolved short life spans and low fertility. I was saddened but not surprised to hear that she had trouble getting her results published, simply because they went against the established dogma.
Clearly, what Gruber observed is an outcome that is adaptive for stabilizing the community of rotifers. But dogma says that evolution always seeks to maximize the reproduction of the individual, no matter what the consequence for the community. Hence the official skepticism of her results: if the rotifers harbor this capacity to both to produce more eggs and to live longer, then what could keep that trait from quickly rising to dominance? (The answer, of course, is group selection; but peer review is often influenced by gatekeepers who deny the reality of group selection.)
What is a rotifer?
Rotifers, it turns out, are all around us. They occupy that size regime (along with mites and nematodes) that so frequently escapes our attention: much larger than single-cell species, but still too small to see. The oceans, every pond, every stream and many puddles are full of rotifers. Even mossy patches in a wet forest carry rotifer populations. Wherever there is water, they thrive; and where water is intermittent, they go into a state of suspended animation, waiting for the next rain. There are 2,200 known species of rotifer, and counting. The largest are about 2mm long, the smallest are but a speck to the eye.
Rotifers are an important part of the freshwater zooplankton, being a major foodsource and with many species also contributing to the decomposition of soil organic matter.[ref]
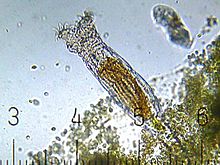
photomicrograph from Wikipedia’s article on rotifers
At banquet dinner, Gruber filled me in on the sexual versatilty of rotifers. When conditions are stable, they just clone themselves. They lay eggs that are exact copies of the mother. No sex.
Under stress, mother rotifers lay eggs that can also develop into females. Females lay eggs that develop exclusively into males in the next generation. Some of these are males, and they can mate with their own mothers or others of her generation; in this case, the eggs produced will always become female.
To summarize: the parthenogenic form reproduces more parthenogenicists or females. Females can reproduce as males with the exact same genome as the female. Female + male can combine to produce another female, with genes that derive half from each parent.
Gruber also told me that a male rotifer is not so impressive a specimen as a bull or a peacock. In fact, males are tiny tiny, only about 1/10 the size of the female of the same species. Males cannot eat or grow. They live for just one thing, and they don’t live very long. They latch onto the female body and inject their sperm in any place that happens to be near at hand. It doesn’t seem to matter; the sperms navigate through the female body, and find their way to the ovaries.
There are more curiosities and mysteries associated with rotifers. The bdelloid family of rotifers have no sex at all, and have not known sex for at least tens of millions of years. Woody Allen asks why they bother to get up in the morning. Evolutionists ask, how do they manage to keep their genomes from succumbing to inexorable accumulation of deleterious mutations. (This is Mueller’s Ratchet).
And rotifers are usually found in extended colonies. What do they get from one another?
Keynote by Austad
Closing the conference was a keynote address by Steven Austad. Austad gets along with everybody. He is widely knowledgable, and famous for his radical common sense. He described his induction into the field, as a grad student in the 1970s. The central dogma of his time was that aging could be observed only in protected environments like a zoo, but that animals in nature died of other causes before they could die of old age. But, working with an island population of opossums as a young student, Austad captured many that were old, some that were clearly very impaired and not long for this world, yet still reproducing.

Austad warned us that much of what we have long assumed about the biology of aging is not to be taken literally without exception; and some of it is merely persistence of myth.

Life spans of mammals (y axis) vs body mass (x axis) in a log-log plot
He showed us the classic log-log plot of animal size vs lifespan. In mammals, life span rises slowly, with about the 1/4 power of an animal’s weight, which corresponds to a slope of 0.25 in the log plot. There are outliers where animals have managed to find strategies to suppress their death rates from predators and disease. Most birds live longer than comparably-sized mammals, and the most dramatic examples are people and bats.
I had known that mice are outliers on the downside. Since mice provide food for a great number of predators, and they freeze to death over the winter; their life spans are below the trend line. What I learned from Austad is that the exceptions extend to all small rodents. For rodents less than 8 kg, there is no correlation at all between size and life span. No one, to my knowledge, has explained this.

The hard thing for me to hear was that, as a way to extend life, caloric restriction is far from perfectly robust and universal. He reminded me of an experiment a few years ago with 41 diverse strains of out-crossed mice. The mice were “recombinant inbred” = first generation crosses between different purebred strains. Under 40% caloric restriction, about a third of these showed life extension, a third showed no significant life extension, and a third actually lived shorter when restricted. There were more mice with shorter life spans under CR than with longer life spans!
Finally, strain-specific lifespans under CR and AL feeding were not correlated, indicating that the genetic determinants of lifespan under these two conditions differ. These results demonstrate that the lifespan response to a single level of CR exhibits wide variation amenable to genetic analysis. They also show that CR can shorten lifespan in inbred mice…
Strikingly, the majority of strains showed no extension of lifespan under the level of DR used in this study (Figs. 1C, D). Only 5% of the strains for males and 21% of the strains for females showed statistically significant life extension under DR (p values < 0.05)…
Of note, the longest lifespans achieved under DR did not exceed the longest achieved under AL feeding. [ref]

Differences in life span between CR and full-feed, male (left) and female (right) strains.
Austad himself did a study of CR for mice captured from the wild.
Although hormonal changes, specifically an increase in corticosterone and decrease in testosterone, mimicked those seen in laboratory-adapted rodents, we found no difference in mean longevity between ad libitum (AL) and CR dietary groups. [There was] higher mortality in CR animals early in life, but lower mortality late in life. A subset of animals may have exhibited the standard demographic response to CR in that the longestlived 8.1% of our animals were all from the CR group. Despite the lack of a robust mean longevity difference between groups, we did note a strong anticancer effect of CR as seen in laboratory rodents
This study demonstrated an increase in maximum life span, but not mean life span under CR. Many people in life extension are very interested in extension of maximum life span, because, as they say, it demonstrates that the fundamental biology of aging has been affected. I agree, but note that increase in maximum without mean life span is the nightmare we have here. It means that the biology of aging has been affected, but not in the same direction for everyone. There are just as many mice in this study whose life span is shortened by CR as the ones whose life span is lengthened. This is what I find most troubling. I once had a graduate advisor who nailed a particular human tendency when we relate ourselves to what we know about others: “Statistics are for everyone else; dumb luck for me.”
As in his past work, Austad offers so much useful good sense in his keynote…And yet he clings to a view that aging is driven by an accumulation of damage, that it can be slowed but never reversed, that there are no genetic mechanisms that have evolved solely for the purpose of assuring a fixed (shorter) life span. The three points are related but not identical. Curiously the idea that damage is the root of aging is not the influence of evolutionary theorists, but far older, rooted in ancient concepts of impermanence.
I have written an academic article [ref] and two blog posts [one, two] in opposition to “wear and tear” theories, and devoted a chapter of my forthcoming book to the subject. It is the most common misconception in the field that aging in biological organisms is akin to physical wear and chemical entropy, and that it has something to do with the Second Law of Thermodynamics.
I know it is theoretically possible, and hope that it will prove generally true in practice, that the body knows how to repair all the important kinds of damage that accrue in aging, and is capable of restoring itself to a youthful state, given the appropriate signaling environment.
Austad’s present research is based on the observation that misfolded proteins tend to accumulate in our cells, and are related to dysfunction and disease, most prominently Alzheimer’s. Long-lived varieties need to keep proteins in the right conformation, with “chaperone” molecules that are particularly effective. Austad is isolating and transplanting some of these chaperone molecules from his menagerie of 500-year-old clams.
Despite differences in theoretical perspective, I have found the community of aging biologists to be especially personable and gracious. I have known Austad and Kirkwood in the deep past, and Baudisch more recently because she belongs to the next generation. Before I had any reputation or credibility in the field, all of them responded to me personally and respectfully. The most promising thing to come out of the meeting for me personally is that I told Austad privately of my idea to test hundreds of combinations of life extension treatments, in order to learn how they interact (see my blog from last month). He told me about an NIA program that evaluates proposals for experiments with mice, deadline later this year. The program is not terribly oversubscribed because it offers no funding to the winning proposals; however winning proposals will be assigned each to three separate mouse labs around the country that will replicate the experimental design in triplicate. I’m pumped!
Privately, Austad also told me that a previous winner had proposed combining rapamycin with metformin and the test was successful. In yet unpublished results from three labs, the combination of metformin and rapamycin extends mouse life span more than the sum of the benefits from the two separate treatments.
Thanks for the report!
Typo in “Aging Is Not a Process of Wear and Tear”:
Loose analogies may suggest that at some point in the life
of an organism, damage accumulates to the point where
repair of the organism is energetically _less_ costly than replacement
through reproduction. This idea is implicit in the
foundation of the Disposable Soma theory.
I suggest you mean “more” where “less” appears above.
Typo in “Aging Is Not a Process of Wear and Tear”:
Circling for the wounded beast, LaPointe and Hekimi …
Suggest you want:
Circling the wounded beast, LaPointe and Hekimi …
> “[…] a mathematical science that yields specific and quantitative information […]”
heh, a tautological fallacy appealing to authority. (the Queen of Sciences in this case)
Cellular automata is no less mathematical (in fact orders of magnitude more formal) and yet far from predictable in less than trivial cases. Whenever feedback loops exists, the human mind exposes its inadequacy. (e.g. 3 body problem)
> ” multifaceted, ”
and by multifaceted you probably mean a multidimensional pareto optimal surface on every variable that may affect it. 🙂
Good luck with the proposal, Josh. I predict success.
Thanks Josh, your report corroborated what I thought I knew. Most interesting to me was the mitochondrial flashes- the rate flashing at the early age peak seems to dictate the speed at which subsequent life plays out. That implies both that aging is programmed, and that subsidiary clocks controlling the ‘playing out’ of the processes, are ‘set’ at that point. Of course that’s a clue to resetting them.
Anyone who knows me as Josh certainly does, know that I despise the notion that evolution is a mathematically precise science with any predictive power – at best evolutionary theory is incomplete (for the reasons Josh mentioned and more), at the worst – and I believe this to be the case, it’s wrong and leading us to wrong conclusions.
Thanks a very interessting report.
Quick vote. Do you then all skip CR/IF?
Personally, I’ll keep my CR/IF regimen, as it gives me more energy. It takes more than a single study on mice to make me quit CR/IF.
Personally, I’m with you. I keep my body lean by not eating starches, by fasting a day a week, and abundance of exercise of all sorts. It works for me.
If you ask me, I say to experiment with your body and see what works for you. I think you can feel what’s good for you, especially if you develop the sensitivity.
What about the hypothesis that it is not caloric restriction but protein or more specifically methione restriction that works in humans as proposed at the website nutritionfacts.org of Dr. Michael Greger, a physician which I highly respect because his website is full of very insightfull information based on scientific evidence. See Methionine Restriction as a Life Extension Strategy at his website : http://nutritionfacts.org/video/methionine-restriction-as-a-life-extension-strategy
I would be glad if you could comment about this strategy.
I think there is good evidence that methionine restriction slows aging. I don’t think it’s a practical strategy for humans because (1) animals on methionine restriction crave methionine; and (2) diets without methionine must be chemically reconstructed from protein constituents. There is no way to adequately restrict methionine with a diet based in any ordinary foods, even vegan ones.
Could one at least argue that vegan food although not perfect does significantly slow aging because it contains much less methionine than animal proteing?
There are good reasons for thinking a vegan diet leads to longevity. But I believe that methionine restriction does not work linearly (the less methionine the better) but rather it kicks in only at very low methionine levels which are impossible to achieve in practice without synthetic foods.
I think it may be possible that where CR reduces life span of mouse, its because it started too early. I wonder it that meta study took into consideration when the mice were put on CR.The paper says mice were put on AL or CR feeding from 2-5 months.
However this Longo paper cites research with lifespan extending benefits of DR started at 12 months.
http://diyhpl.us/~nmz787/pdf/Fasting_vs_dietary_restriction_in_cellular_protection_and_cancer_treatment_from_model_organisms_to_patients.pdf
And prolonged fasting is by far less studied than CR proper. Makes it lot more risky though, too.
The theory of Mitoflashes has been heavily criticized in Nature
“The interpretation of mitoflashes as ‘bursts of superoxide radicals’ assumes that circularly permuted yellow fluorescent protein (cpYFP) is a reliable indicator of mitochondrial superoxide2, but this interpretation has been criticized because experiments and theoretical considerations both show that changes in cpYFP fluorescence are due to alterations in pH, not superoxide3, 4, 5, 6, 7. Here we show that purified cpYFP is completely unresponsive to superoxide, and that mitoflashes do not reflect superoxide generation or provide a link between mitochondrial free radical dynamics and lifespan. ”
http://www.nature.com/nature/journal/v514/n7523/full/nature13858.html
Yes, I’m aware of this other interpretation, and Dr Dong spoke with me about it, and why she thinks this is a marker for superoxide and not for pH. I don’t have the background or the expertise to take a side in this question.
Here is her rebuttal: http://www.nature.com/nature/journal/v514/n7523/abs/nature13859.html
INteresting article about bone and muscle loss in aging mice.
http://europepmc.org/abstract/med/16750436
Leptin levels and inflammation is correlated with muscle and bone loss.
Whats interesting is that body mass and bone keeps growing until 12 months.
I always wonder how to translate mouse aging to human aging. Are mouse like people having a compressed childhood or like people getting sexually mature at kindergarten age?
Great article, Josh – and I’m really interested in the rotifer experiments. However, is group selection actually sufficient to explain the results? If group selection were the dominant force, I would expect some low food tanks to feature population booms and crashes as long-lived, high-fertility animals took over the population and then all starved. The end result would then be that, averaging across all low food tanks, the short-lived low-fertility rotifers made up the majority of the low-food population.
It doesn’t seem like the results you describe rule out a decrease in lifespan and fertility simply due to sickliness caused by malnutrition. But I understand you can’t fit a complete description of Gruber’s work into a short article, so maybe I’m missing some important context.
Gruber’s work is yet unpublished, but she promised to send me a preprint when she has written it up and submitted it.
In the meantime, my guess is that the “group selection” took place a long time ago, certainly not in her experiment.
Another interesting experiment with FGF21 transgenic mouse. FGF21 is upregulated in starvation.
http://elifesciences.org/content/elife/1/e00065.full.pdf
30-40% median lifespan extension.
That was an very interesting experiment. A methionine restricted diet increases FGF-21 by 5 fold in 2 month old mice and 2.5 fold in 12 month old mice.
Methionine restriction restores a younger metabolic phenotype in adult mice with alterations in fibroblast growth factor 21
http://www.ncbi.nlm.nih.gov/pubmed/24935677