
The Vital Question, by Nick Lane, 352 pp, Profile Books, April, 2015
Life has been on earth about 4 billion years. If we think of each billion as a term at college, then for its Freshman, Sophomore and Junior years, life majored in chemistry. Every possible chemical environment was probed, drawing its first energy from the warm, hydrothermal vents at the bottom of the ocean, leaving the alkaline silos where life got its start, colonizing the sea and the land, the atmosphere, places cold and hot, wet and dry, acid and base, high in the clouds and deep in rock miles under the earth.
Life as an underclassman was just one cell at a time, and a small cell at that, maybe a micron across, a “prokaryote” that despite its impressive chemical virtuosity had little physical structure and the simplest life cycle. Divide and conquer.
Before his Sophomore year was over, the precocious chemists had figured out how to use the sun’s energy to pull carbon right out of the air, and the entire atmosphere had been processed, molecule by molecule, from (inert) CO2 to (energy-laden) Oxygen. The energy economy of the earth was transformed.
Then, about a billion years ago—life was a rising Senior—a once-in-a-lifetime event occurred, a wild fluke. One of these chemists that specialized in membranes and electrochemistry was invaded and colonized by a parasite that specialized in combustion chemistry—oxidation of sugar, to be specific. The invader put all that atmospheric oxygen to good use, then spewed out toxic ROS (reactive oxygen species) that almost killed the host, as it had killed many archaeons before. But this time, the host survived and, over an amazingly short time of just 2 million years, learned not just how to coexist with the invader, but also to domesticate the parasite and put him to work. The host was already using ATP as an energy source, and the parasite had a talent for producing copious quantities of ATP/energy—more than any archaeon had ever seen before. Stick with me, kid and we’ll go far. By the end of Senior year, every plant and animal on earth, every fungus and amoeba, toadstool, jellyfish and (by the way) you and me—every cell in our bodies, all of them are descended from that one sick, infected “hopeful monster”. Her name was LECA, the Last Eukaryotic Common Ancestor. Everything we think of as macroscopic biology comprises her progeny, in so many different guises.
Because it was in its Senior year that life first got interested in engineering, cell upon cell. Pipelines and networks of roadways, electrical circuits, information processing within and between cells. Muscles, bones, shells, levers, motors and other mechanical devices that were made out of living cells! Locomotion in the sea, on land and air. The greening of earth and sea, predators and prey, webs of interconnected life—all of this is the realm of eukaryotes.
This story wasn’t written by Nick Lane, but he certainly tells it with a flare. The story was written by biological visionaries of the last century. Richard Goldschmidt, Francis Crick, Lynn Margulis, Carl Woese are biologists that I know. Also Stephen Jay Gould and E. O. Wilson—I’m sure there are others who are equally eminent with whom I am unfamiliar. Erwin Schroedinger and Freeman Dyson were physicists who also contributed to the canon, speculating about the Great Questions.
In our generation, Nick Lane is the only one I know who is making a bid as heir to these giants. (I hope that readers of this column will use the comments section to make me aware of others.) He is a biological visionary who happens to be a great storyteller. In a series of books since 2005, he is asking the Great Questions about life, how it began, why it is the way it is, how much of life’s history is pure chance, and if we should run into an extraterrestrial life form, how much can we expect it to be like us? His latest book is called The Vital Question. Like his previous books, this one takes a bioenergetic view of life. The centerpiece of the present volume is that singular event at the beginning of Senior year: The fusion of (probably) several bacterial and archaeon individuals to form that one “hopeful monster”, the mother of us all.
Among the traits that all eukaryotes have in common (but distinct from prokaryotes) are
- a cell nucleus, with multiple linear chromosomes. (Prokaryotes, by contrast, have their DNA in a single loop and many smaller loops.) “The nucleus is an exquisitely adapted structure, no mere repository for DNA.”
- genes coded piecemeal, interrupted by introns, a catch-all word for the 95% of DNA that does not code for a protein, and may have many different functions, or possibly none at all
- within-species genetic exchange (sex) with crossover of parts of chromosomes. Before eukaryotes, genetic exchange was rampant, promiscuous, and willy-nilly. Every type of bacteria exchanges genes with every other type.
- energy generation in mitochondria (the parasites mentioned above)
- endoplasmic reticulum, a network of membranes that guides transportation of molecules within the cell
- Golgi apparatus, there are many within the cell, and they serve as post offices, addressing and dispatching packages or protein to their appropriate destination
- soft, permeable cell membranes that can take in selective nutrients or even engulf and absorb another cell in its entirety (phagocytosis).
Eukaryotes are about 100,000 times bigger than prokaryotes, and there is much more structure and machinery. Right from the start, Lane frames his narrative with a question about eukaryotes, why they are so different from everything that came before, why they were able to do things that in 3 billion years the prokaryotes never attempted, and why there are no extant “missing links” by which we might trace the evolution from prokaryote to eukaryote.
There is a black hole at the heart of biology. Bluntly put, we do not know why life is the way it is. All complex life on earth shares a common ancestor, a cell that arose from simple bacterial progenitors on just one occasion in 4 billion years. Was this a freak accident, or did other ‘experiments’ in the evolution of complexity fail? We don’t know. We do know that this common ancestor was already a very complex cell. It had more or less the same sophistication as one of your cells, and it passed this great complexity on not just to you and me but to all its descendants, from trees to bees. I challenge you to look at one of your own cells down a microscope and distinguish it from the cells of a mushroom. They are practically identical. I don’t live much like a mushroom, so why are my cells so similar? It’s not just that they look alike. All complex life shares an astonishing catalogue of elaborate traits, from sex to cell suicide to senescence, none of which is seen in a comparable form in bacteria. There is no agreement about why so many unique traits accumulated in that single ancestor, or why none of them shows any sign of evolving independently in bacteria. Why, if all of these traits arose by natural selection, in which each step offers some small advantage, did equivalent traits not arise on other occasions in various bacterial groups?
This story, the merging of very different life forms to create evolutionary revolutions, is associated most closely with Lynn Margulis. Lane acknowledges the deep legacy of Margulis, and also parts company with her and diverges from her Story of Life to offer his own version.
Something odd seems to have happened at the very origin of eukaryotes. It looks like the first eukaryotes picked up thousands of genes from prokaryotes, but then ceased to ply any trade in prokaryotic genes. The simplest explanation for this picture is not bacterial-style lateral gene transfer, but eukaryotic-style endosymbiosis. This is the simplest possible scenario for the origin of eukaryotes: there was a single chimeric event between an archaeal host cell and a bacterial endosymbiont.
Demise of the the Tree of Life
To understand the oddness, we go back to the “tree of life” that was Darwin’s inspiration.
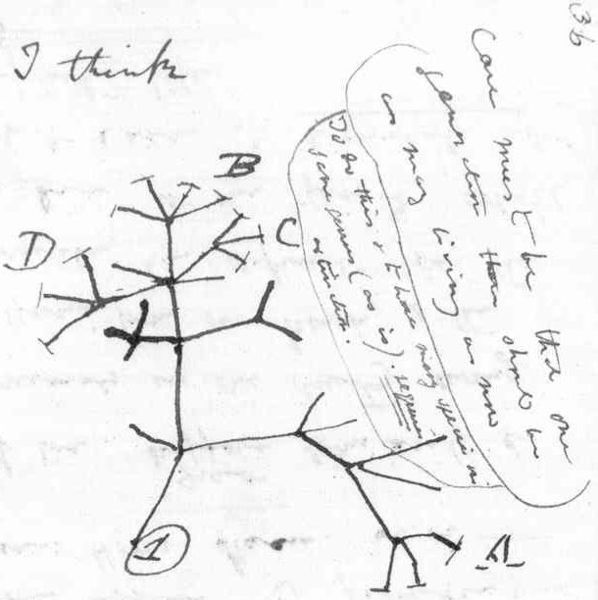{kind=link}

Through evolutionary history, species diverged via mutations and split off from one another, so that one species became two, in a branching process. In this picture, due to Darwin, relatedness is perfectly defined, and every species has a unique path tracing it back to LECA.
Once genome analysis became possible in the 1980s, it was an early project to try to reverse-engineer the tree. But the result of that project was a huge surprise. Tracing different genes produced different trees, until it became clear that there was no tree at all, but rather a web of interconnecttions. Everywhere there were cross-links in the tree. Every individual species had acquired genes from many different places.

How can we reconcile this with the common fact of our experience that every cell comes from a single parent that divided in two? (Yes, there is sexual combination, so a cell can have two parents, but in our experience these two parents are always very closely related, in fact they are by definition of the same species.)
And most problematic of all is the genome of LECA, parts of the genome that all eukaryotes share. Those genes seem to have come from a dizzying array of very different bacteria and archaeons. How can that be reconciled with the idea that LECA arose in a single, improbable event? How can there be both a unique LECA and such a diverse genomic ancestry?
Different genes in the same eukaryotic organism do not all share the same ancestor. Around three-quarters of eukaryotic genes that have prokaryotic homologues apparently have bacterial ancestry, whereas the remaining quarter seem to derive from archaea. That’s true of humans, but we are not alone. Yeasts are remarkably similar; so too are fruit flies, sea urchins and cycads. At the level of our genomes, it seems that all eukaryotes are monstrous chimeras. That much is incontestable. What it means is bitterly contested.
It was this diversity of genes from many origins that Margulis cited as strong support for her thesis that merging of different, unrelated species has time and again seeded evolutionary saltations (times of abrupt change). But Lane has a different idea. These diverse genes were acquired all at once during a brief period (~2 million years is Lane’s astonishing claim) around the time of LECA. The reason that they now appear to be associated with diverse kinds of bacteria is that bacterial genomes are easily mutable via the bacterial brand of promiscuous sex, the exchange of plasmids. The apparent diversity of bacterial sources for eukaryotic genes is an illusion based on the taxonomic groupings (“species”) of bacteria today, which may be very different from their groupings of old because genomes are continually reshuffled across the diversity of different bacterial “species”.
Neither Margulis’s story nor Lane’s adequately addresses the greatest mystery (as Lane himself is first to admit): why there are no surviving missing links between the prokaryotes and the eukaryotes?
The Energy Metabolism and the Origin of Life
Lane’s perspective centers on life’s capacity to capture energy and use it for purposes internal and external. His singular contribution from the past was to remark how strange and curious is life’s universal energy factory: energy is stored and harnessed as a potential difference across an organic membrane. So it is with all life’s diverse forms adapted to diverse environments. It was the central insight of Lane’s 2005 book Power, Sex, Suicide that this may be a hint as to how life formed. There are geothermal vents underwater, where metal oxides are spewed copiously into the sea, creating an alkaline pocket in an acid ocean. Here are also mineral structures with micro-pores of cellular dimension. So two of life’s necessities were freely available as foundation for pre-biotic evolution: compartmentalization (micropores) and energy in an appropriate form (gradient of H+ ion concentration). Lane summarizes and elaborates this story in the new book.
Eukaryotes arose when mitochondria first appeared as an endosymbiont in LECA’s interior, providing a generous and reliable source of energy in the form of ion gradients, with input from biotic fuel and ambient oxygen. It is Lane’s ambition to explain the broad outlines of eukaryotic life from this one event—the diversity, the behaviors, and the similarities of all plant and animal life, all derived from the character and circumstances of mitochondria.
It means, astonishingly, that mitochondrial variation alone can explain the evolution of multicellular organisms that have anisogamy (sperm and eggs), uniparental inheritance, and a germline, in which female germ cells are sequestered early in development—which together form the basis for all sexual differences between males and females. In other words, the inheritance of mitochondria can account for most of the real physical differences between the two sexes.
Lane even takes a stab at explaining the life cycle, including aging. I applaud his vision and ambition in stepping back to look at the big picture and addressing the Great Questions. To what extent are his answers convincing? I’ll continue next week with some ways in which Lane’s perspective offers fresh new understanding, and some equally Great Questions that he does not address.
Hello Josh,
I’ve been enjoying your Aging posts for some time now. I just read the review of Nick Lane’s book, and it prompts a question.
I am considering writing something on “chance.” Your opening paragraphs on the beginnings of life made me wonder if you could address this:
Can one speculate how unlikely it was that one “chemist” was invaded by just the right specialized parasite? Might it never have happened?
Thanks for taking the question.
Best,
Gus Russo
Gus –
I don’t know what you mean “might it never have happened?” We’re pretty sure it did happen. Do you mean, ‘was it just a lucky break?’ Maybe there are a million other planets where life evolved and got as far as bacteria, but stopped there because they didn’t get that lucky break.
The answer is, I don’t know. And I’m pretty skeptical of anyone who claims he does know. The probabilities involved in evolution cannot be computed or even estimated from first principles.
A lot of people, scientists and lay people alike, tend to think “it must have had a high likelihood of happening, because it did happen.” But of course they’re wrong, and we don’t know that. It may have happened just once in a billion planets in each of a billion galaxies in each of a billion universes, but of course that’s the one we’re looking at because we are its product. If we had contact with any extra-terrestrial life form, that would change all the probability judgments immediately, but without that, we don’t know whether, given a billion years, the likelihood of an earth-like planet evolving life is 1 in 2 or 1 in a googol (10^100).
The first eukaryote looks like a huge leap, difficult to explain. But a lot of other things that evolved also look like a huge leap. I think maybe there’s something we don’t yet understand, something that ups the probabilities and makes all these developments plausible. Maybe it has something to do with evolution of evolvability. Maybe it has to do with quantum evolution.
– Josh
Hi Josh, “All life starts and ends with an energy gradient.” You might enjoy the books by David Valentine on DHA/EPA and membrane dynamics viz a viz the “origin of species”! Best, Fred
This volume stands out from the crowd by underlining crucial aspects of biology which, although well evidenced are not yet generally recognized. Not only by the layman, to whom the work is primarily addressed, but also by the majority of individuals working within the sciences, even areas of biology.
The first, which in the light of present knowledge should be virtually self-evident, is that there can be no firm borderline to be drawn between chemistry and biology. That they are are entirely contiguous. Indeed, that biology is truly a subset of chemistry. And the underlying energetics are the same.
Of course, the established biological phase has its own set of characteristics, proactive feedback mechanisms and self-replication being among the most noteworthy. But they are simply different kinds of chemistry.
Lane does diverge from the interpretation presented in my own writings inasmuch as he seems to be reluctant to break with the tradition of equating biology to “life’. The difference is mainly semantic, but I feel it is more useful to identify “life” with the increase of intricacy of our universe that can be traced at least as far back as the assembly of the chemical elements in the first stars. An evolutionary continuum that has its latest manifestation in the emergence of technology and its latest spearhead, the Internet.
The second salient issue addressed is that at last we can construct a plausible model of abiogenesis drawing upon our relatively new knowledge of sub-oceanic hydrothermal vents and the suggestions arising therefrom, notably by Peter Mitchell.
The serious weaknesses in the “primeval soup” and other previous models were nicely addressed by Robert Shapiro in his 1986 book “Origins”. They are simply not consistent with well established principles of chemistry and physics. Similar implausibilities arise with “DNA first”, “RNA first” “protein first” and “cell first” models. Now, within the context of the microcavities of the alkaline vent plumes, we have a situation in which the energetics, and the chemistries are favorable. Furthermore, we have innumerable matrices which the generation of cell membranes can plausibly occur. In my latest book “The Intricacy Generator: Pushing Chemistry and Geometry Uphill”, I sketch out a scenario in which co-evolution of cell membrane, protein, and nucleotides could take place with acceptable probability.
Thirdly, with regard to biology that might occur on other worlds, Nick lane astutely points out that:
“It is going to be carbon-based, it is going to be driven by this peculiar mechanism of energy generation – protons across membranes,It is going to have the same structural constraints, it is going to have to deal with populations of cells within cells.”
One area where I find Lane’s arguments rather less convincing is in his contention that the advent of eukaryosis was a singular event. Which is not consistent with the usual incremental and largely stochastic mechanisms that characterize evolutionary processes. Maybe that’s simply my reluctance to believe in “miracles”. But lending some weight to my skepticism on this point are genome studies of the deep sea archeon Lokiarchaeota which are suggestive of intermediates.
Furthermore an even stronger clue to the transformation from prokaryote to eukaryote has been found by Masashi Yamaguchi et al. In this case, the fortuitous discovery of an extant deep sea organism (Parakyron myogenensis), the morphology of which suggests a transitional entity.
“The Vital Question” is a “must read” for all who have any interest in these matters and would also make a very worthwhile ancillary to any secondary school curriculum.
Does it mean that an eukaryotic cell contains a lot more energy for animals that live as filter feeders?
If you are interested in other scientists working with biology, could physicist Nigel Goldenfeld offer something?
https://www.quantamagazine.org/seeing-emergent-physics-behind-evolution-20170831/
Some quotes:
“The physicist Nigel Goldenfeld hates biology — “at least the way it was presented to me” when he was in school, he said. “It seemed to be a disconnected collection of facts. There was very little quantitation.” That sentiment may come as a surprise to anyone who glances over the myriad projects Goldenfeld’s lab is working on. He and his colleagues monitor the individual and swarm behaviors of honeybees, analyze biofilms, watch genes jump, assess diversity in ecosystems and probe the ecology of microbiomes. Goldenfeld himself is director of the NASA Astrobiology Institute for Universal Biology, and he spends most of his time not in the physics department at the University of Illinois but in his biology lab on the Urbana-Champaign campus.”
“In the earlier phase of life, even the core machinery of the cell was transmitted horizontally. Life early on would have been a collective state, more of a community held together by gene exchange than simply the sum of a collection of individuals.”
“Tommaso Biancalani and I discovered in the last year or so that life automatically shuts off the horizontal gene transfer once it has evolved enough complexity. When we simulate it, it basically shuts itself off on its own. It’s still trying to do horizontal gene transfer, but almost nothing sticks. Then the only evolutionary mechanism that dominates is vertical evolution, which was always present.”